Ciclo cellulare
Il ciclo cellulare è l'insieme delle fasi tra due divisioni cellulari consecutive. Il processo inizia quando nasce una nuova cellula, discendente da un'altra cellula che si è divisa, e termina quando quella cellula subisce la divisione successiva e dà origine a un'altra coppia, che sono considerate sue figlie.
Il ciclo cellulare può essere inteso come una serie di eventi che si verificano in modo ordinato mentre una cellula cresce e alla fine si divide in due cellule figlie. Le cellule passano attraverso due fasi: l'interfase (stato di non divisione) e la fase M (stato di divisione).
Nell'interfase, la cellula svolge alcune funzioni specifiche mentre procede verso la divisione cellulare. La fase iniziale è conosciuta come fase G1, quando inizia a sintetizzare RNA e proteine. In questa fase la cellula raggiunge un raddoppio della massa e delle dimensioni. Poi arriva la fase S con la sintesi del DNA e la duplicazione di ogni cromosoma.
Il ciclo cellulare continua con la fase interfase G2: la sintesi dell'RNA e delle proteine continua e inizia la divisione. A questo punto la cellula entra nella seconda fase, chiamata M-phase.
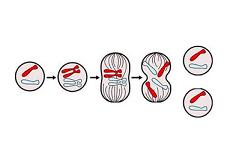
Questa M-phase è quando avviene la divisione cellulare: la cellula madre si divide in altre due cellule (le cellule figlie), che sono identiche. La fase M comprende la mitosi e la citochinesi.
La mitosi è un processo biologico che ha luogo nel nucleo di una cellula eucariotica, appena prima che si divida; in breve, implica che il materiale ereditario caratteristico venga condiviso equamente. La citochinesi, invece, è la divisione fisica del citoplasma in due cellule.
Regolazione del ciclo cellulare
Nel 2001 è emersa una spiegazione della regolazione del ciclo cellulare, che può essere osservata negli organismi eucarioti dal punto di vista delle decisioni prese in alcuni momenti critici del ciclo stesso, in particolare la mitosi. Questo solleva domande come il perché il DNA si replica solo una volta, o perché l'euploidia cellulare può essere mantenuta.
La risposta potrebbe risiedere nel fatto che durante la fase G1 la ciclina facilita l'aggiunta di regolatori chiamati Cdc6 al complesso di riconoscimento dell'origine (ORC), che è responsabile di sollecitare il macchinario di replicazione genica in un processo in cui si genera un complesso per la futura replicazione del DNA.
All'inizio della fase S, Cdk-S genera la dissociazione di Cdc6 e degrada le sue proteine, ed esporta Mcm nel citosol, in modo che non sia possibile per l'origine di replicazione reclutare un complesso pre-replicazione fino al ciclo successivo. Per tutta la fase G2 e M, l'unicità di questa struttura è mantenuta fino a quando il livello di attività Cdk diminuisce dopo la mitosi e diventa di nuovo possibile aggiungere Mdm e Cdc6 per il ciclo successivo.
Un'altra questione sollevata da questo studio è come si entra nella mitosi. Per rispondere a questo possiamo pensare che la ciclina B, che è comune nella Cdk-M, sia presente durante tutto il ciclo. La ciclina è solitamente inibita dalla fosforilazione attraverso una proteina chiamata Wee; tuttavia, quando la fase G2 si avvicina al completamento, un fosfato chiamato Cdc25 viene attivato e rimuove il fosfato inibitorio per aumentare la sua attività. Attiva anche Cdk-M e inibisce Wee, causando un feedback positivo che porta all'accumulo di Cdk-M.
